International Asclepiad Society
Morphology and pollination of stapeliads
Stapeliads, which are stem succulents, belong to the family Apocynaceae and subfamily Asclepiadoideae. Until recently, they were placed into the Asclepiadaceae family. The stem shapes are very similar in most genera, but when they bloom, the beauty of the flowers is striking as well as their unpleasant smell!
"Stapeliads, Orchids of the Succulent World" is the title of the web pages of late Jerry Barad from New Jersey, USA. The title says everything. The flowers have a beauty and colour that can only be compared with orchids. And they also share another characteristic. The pollen mass is fused in a wax pollen sack - pollinium, which is transferred by pollinators to the style.
All stapeliads have some common morphological characteristics. Succulent stems are the most visible, and which are alike in dif ferent genera. The floral structure of the flow ers is five lobed in all asclepiads and they al so have similar fruits and seeds.
Distribution
Stapeliads are distributed only in the dry regions of the Old World. The easternmost locations are in Myanmar (formerly Burma), Sri Lanka and India and they extend through Pakistan, Iran, the Arabian Peninsula, Israel and northern Africa to the Canary Islands in the west. The northernmost distributions are in Spain and the Italian island Lampedusa and they stretch over northern and north-eastern Africa to southern Africa, where there is the highest density of species. Most stapeliads are terrestrial, hiding in the half shade of grass and bushes. Only some tougher species proudly defy the strong sun.
Stems
The stems are mostly very fleshy. In some species they become woody in the lower part of the stems. They contain a transparent watery sap, which oozes out if injured. Plants are consumed by natives and animals, although the sap is bitter. Stems are angled or round in transverse section. Some species have prominent ribs, most of them have tubercles arranged in ribs. Genera Larryleachia, Lavrania, Pseudolithos, Notechidnopsis and Echidnopsis have very flattened tubercles. In some genera they are conical elongated into a tooth. In Orbea (Orbeopsis, Angolluma, Or beanthus, Pachycymbium) it's one of most distinctive features. The teeth also appear in some species of Huernia and Duvalia (section Arabica). The plants with four ribbed stems are in the majority (Stapelia, Orbea, Huernia, Piaranthus, Carallumaâ¦). In the genera Tavaresia and Hoodia the ribs are more numerous. The above mentioned genera with flattened tubercles have also more ribs, how ever these ribs are not so distinctive.

A - Caralluma (Crenulluma) adenesis, B - Caralluma (Sanguilluma) socotrana, C - Caralluma (Spathulopetalum) gracilipes, D - Duvalia polita, E - Rhytidocaulon macrolobum, F - Rhytidocaulon fulleri, G - Stapelia olivacea, H - Stapelia grandiflora, I - Huernia hybrid, J - Huernia guttata subsp. reticulata, K - Piaranthus geminatus subsp. decorus, L - Pseudolithos migiurtinus, M - Tavaresia barklyi, N - Orbea (Angolluma) sprengeri, O - Orbea variegata, P - Echidnopsis spec., Q - Stapelianthus pilosus, R - Quaqua incarnata var. tentaculata, S - Tromotriche aperta, U - Anomalluma mccoyi, V - Stapeliopsis neronis, W - Hoodia spec. MST 13, X - Pectinaria longipes, Y - Larryleachia marlothii.
Leaves
Only Frerea indica in the whole group of stapeliads has fully developed leaves that fall off at the end of growing season. In other species the leaves are strongly reduced. The leaves in some species in Echidnopsis, Stapelia and Caralluma are like scales that soon dry. They stay in some species and fall off in others. In Huernia pillansii and Stapelianthus pilosus the leaves are transformed into soft hairy tips. The leaves can be transmuted into hard thorns, the most distinctive being Quaqua mammillaris. Leaf thorns are distinctive of the genera Hoodia, Tavaresia and Edithcolea. Mostly, the leaf spine is single, only in the genus Tavaresia a couple of side bristles (stipules) appear beside the central one. Other stapeliads lack the stipules. In some species (Duvalia polita, Orbea (Orbeopsis) lutea only their remains denticles are still visible.


Growth
The growth pattern of stapeliads is very simi lar. Side shoots spring up from the base of the primary stem and they grow further from the lower part of the stems. Some side shoots root in contact with the ground and become inde pendent of the primary root system. They can form large pillows. Older stems in the middle of the clumps die away in time, and the outer stems spread forward and make separate plants. Most of the stapeliads are of low growth only some species of the genus Hoodia can reach one meter and a half. The basic growth form of stems is erect, but in some species, it is creeping (Duvalia, Echidnopsis, Huernia). Some species of genera (Orbea, Boucerosia, Duvalia) form underground stems rhizomes.
Flower formation
Flowers appear in large numbers and can form inflorescences, only rarely do they appear in dividually. They are usually at the apex of the stems. In most stapeliads it seems that the flowers appear laterally to the growing axis or even from the base of the stems (Huernia), however it is not so. From the base of the stems grow a peduncle, and from it the main stem grows out and pushes the peduncle in a lateral position. The peduncle is always between the axis of leaves or tubercles. In some species flowers appear in succession from the same peduncle. In the genera Caralluma, Spathulopetalum, Cryptolluma (by Plowes) the stems elongate to a long, thin peduncle with numerous little flow ers. When the fruits ripen, the whole stem with peduncle dries up. Boucerosia, Apteranthes, Sulcolluma, Crenulluma (by Plowes) have flowers in umbels at the apex of the stems. It moves later to a lateral po sition because of the growth of the stems.

Flower structure

At first sight the flowers are very uniform. They have five sepals and five fleshy petals corolla lobes. The sepals are tiny, the petals are usually in a star formation and grown together to form a shorter or longer tube, at least to one third or nearly for the whole length in some species (Echidnopsis). It these cases, there is only a small slit at the top of the flower to allow pollinating insects to enter. In some others (Ophionella arcuata) the corolla lobes are joined together at the tips and the shape of the flower is lantern like. The corolla in Tavaresia is tubular and trumpet-shaped. The corolla in some species also has a fold in the middle of the flower that forms a ring around the tube and it is called an annulus. In the latest research (Liede & Kunze 1993) the annulus has been recognized as a part of the coronal structure (annular corona) The outside of the corolla lacks vivid colours, only in plants with urceolate or lantern-shaped flowers is it more colourful. The inner side of the corolla with many kinds of shapes and patterns is the exact opposite. The surface is of ten wrinkled, or covered with hairs or papillae, transverse furrows or scars. Red, brown, yellow and violet colours with all of their intermediate tones are predominating. The flowers imitate carrion with their smell and colour, and attract pollinating insects with secretions and also hairs. The diameter of the flowers is from a few millimetres to 50 cm in Stapelia gigantea. The stamina and pistil cannot be found in the flower. They are transformed into a complex structure the gynostegium with a single or double corona. Two carpels are hidden inside the gynostegium and each of them forms a separate unilocular ovary united only by the common style. Filaments are joined together into a staminal column which encloses the ovary. The receptive organ for pollen is five slots found in the lateral position of the staminal column between the lobes of the inner corona and are open toward the base and narrow toward the top of the gynostegium. They are named guide rails or anther wings or staminal locks (Barad 1990). The points of the ovaries and staminal column are grown together to a five-lobed style head. They lie directly above the guide rails.

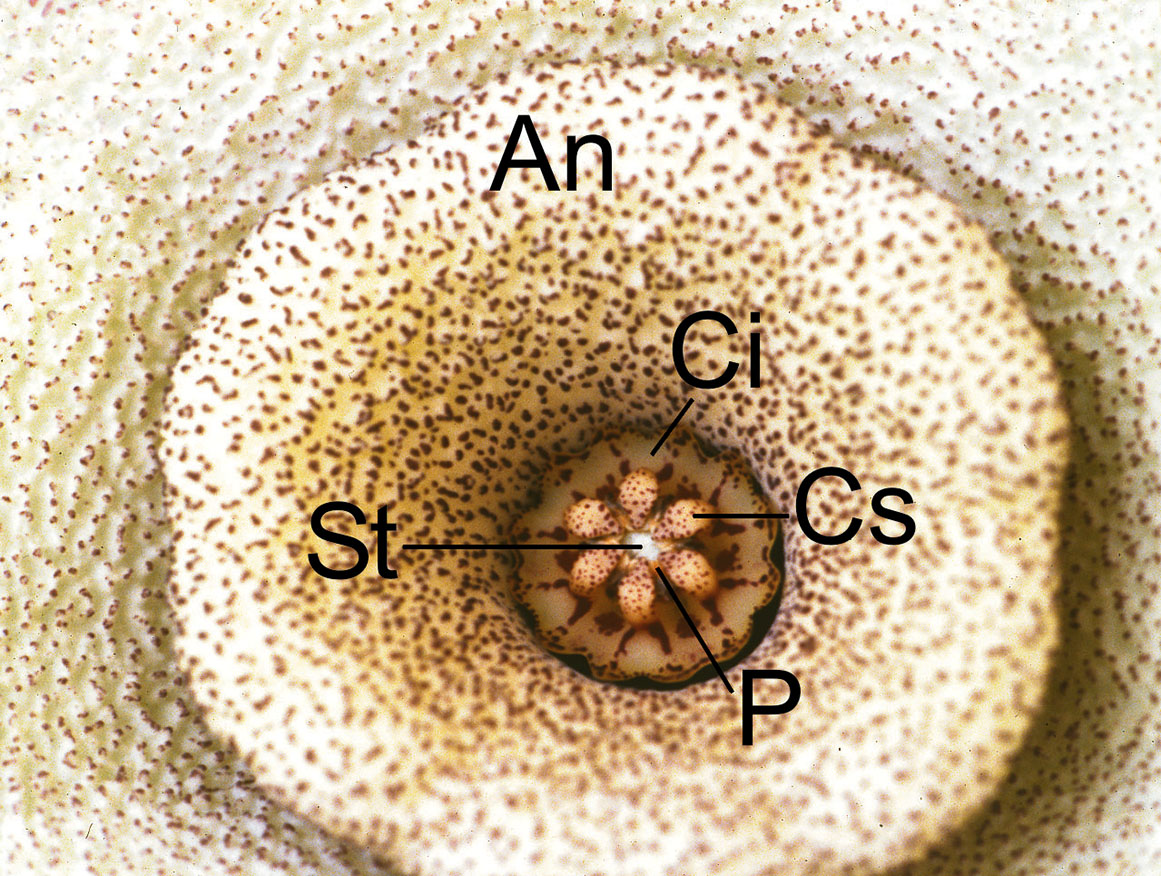

Photo by Matija StrliÄ
The pollen of each staminal area is amassed in two waxy pollen masses or pollinia. A pair of adjoining pollinia is attached to a translator and forming the pollinarium. The translator has a central structure corpusculum (clinging head), a pair of arms of translator (caudicles) and a pair of wings. The corpusculum has a central jag which continues the guide rail. Five pollinaria are positioned in the corners of the style head with the corpusculum directly above the guide rail. The translator is a product of secretion of the underlying glands. The pollinium has a minute wedge-shaped struc ture named the pollinium key (Barad 1990), anchor margin, wave crest or germination crest on the lower inner side. The latter is preferred by most of the authors. The single or dual corona is an appendage to the gynostegium. It consists of the inner or staminal corona and outer or interstaminal corona. Both develop from filaments and anthers. They are five-lobed like other parts of the flower. The staminal corona encloses the staminal column so that the corona lobe is placed between neighbouring guide rails. The corona lobes are usually higher than the style head and they can also cover it. The outer corona can be joined to the base of the floral tube or annulus. It can also be raised and embrace the gynostegium. The interstaminal corona is so reduced in some genera (Piaranthus, Duvaliandra) that it is imperceptible. Nectar cavities are found close under the guide rails. Sometimes they pour out so much sweet liquid that the whole gynostegium is flooded with nectar.
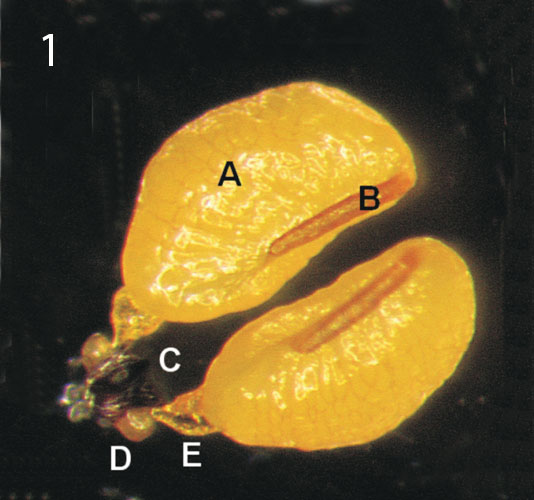
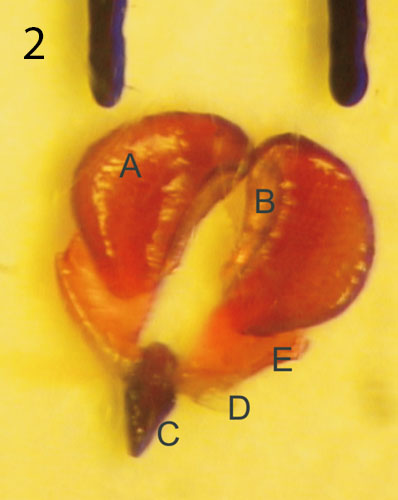

Photo 1 by Matija StrliÄ
Fruit
The corolla with the gynostegium dries and falls away after a successful pollination. Only two naked carpels remain protected with sepals. After successful pollination the pedicel grows fat and sometimes it extends, too. The carpels develop into two fruits follicles which look like horns. It can grow soon after pollination, next spring or even after some years. When the fruit ripens, it bursts longitudinally, and large, up to 7 mm long seeds pop out of it. They are equipped with silky parachutes. The seeds are blown far and wide by the wind, and thus they are distributed far away from the mother plant.


Pollination
Pollination in most of Spermatophyta is performed by the transfer of pollen to the style of the pistil. In stapeliads pollination is more complicated as is the structure of the sexual or gans. The pollinators are usually different kinds of flies, attracted by the odour and colour of the flowers. They look for food in the flowers and lay eggs. The complicated structure of the corona and rigid hairs on the corolla enable flies only a limited access to the nectary glands. They fumble with the proboscis near the slots towards the mouth of the flower. Doing this, the hairs or bristles of the head or legs are often stuck in the guide rails and only one way is possible upwards to the end of the staminal lock, which is connected to the jag of the corpusculum of the pollinarium. If the fly is strong enough, it pulls out the entire pollinarium and moves it to the next flower. If the insects are too weak to remove the pollinarium, they remain trapped in the flower. By searching and drinking nectar in the next flower the pollinium is turned around and the germination crest (pollinium key) turns against the guide rail (staminal lock). Now we see why Barad used the expressions key and lock. The germination crest catches into the guide rail and only one way is possible towards the end of slot where the pollinium finally wedges. The pollinator is trapped again and if it is strong enough, it frees itself or tears off the remnant of the pollinarium at the translator arm. It can pollinate another flower with the remaining pollinium. The placed pollinium remains in the receptive area of the style. Pollen germinates from the germination crest and after a day it is almost impossible to remove it. After a few days the corolla with gynostegium dries up and falls off; only two carpels, which develop into fruit, remain. Some different species of stapeliads usually grow at the same locality, so natural hybridization is possible but not so frequent. Pollinators are usually specialized to one species. The difference in size of the germination crest and guide rails is also a limiting factor.

Photo by Loukie Viljoen

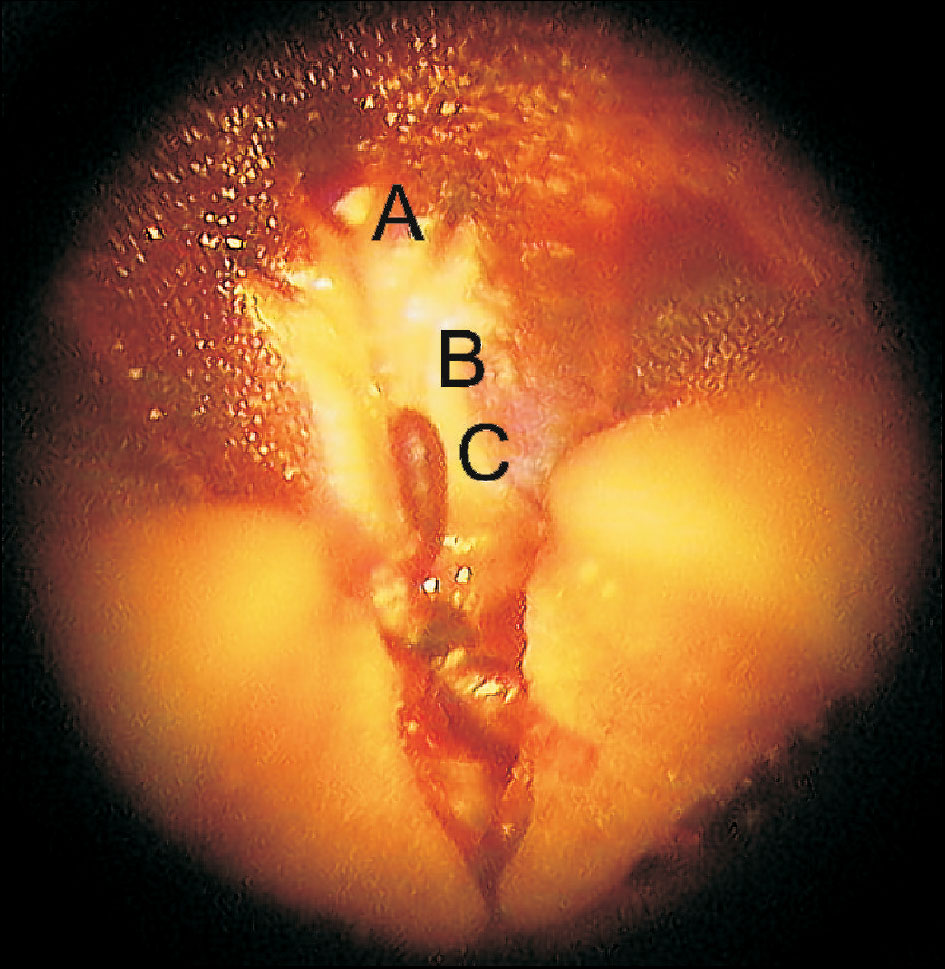
Acknowledgment
I am very grateful to Mr. Loukie Viljoen from Republic of South Africa for allowing me to publish his photo. He also sent me some photographs of a vertical section of Orbea variegata flower that I used to draw the flower vertical section diagram.
About
This article was originally published in Welwitschia 5 (1-4) 2002, journal of Cacti society of Slovenia. English version was published in Asklepios 92, 2005 and German version in Caralluma 2, 2005, journal of IG Ascleps. Text was written by Iztok Mulej. Photos were taken by Iztok Mulej, otherwise indicated.
References:
- Barad, G. S. (1990). Pollination of Stapeliads. Cactus and succulent journal (U. S.), 62 (3): 130-140.
- Liede, S., Kunze, H. (1993). A descriptive system for corona analysis in Asclepiadaceae and Periplocaceae. Pl. Syst. Evol. 185: 275-284.
- Mulej, I. (1994). Nekaj besed o morfologiji stapelijevk. Kaktusi in druge soÄnice 23 (4): 6-9.
- Mulej, I. (1997). OpraÅ¡evanje svilnovk (Asclepiadaceae), Kaktusi in druge soÄnice 26 (3): 4-8.
- Mulej, I. (1998). Pollination of the Asclepiads (Asclepiadaceae), Asklepios 73: 7-11.
- Rauh, W. (1979). Die groÃartige Welt der Sukkulenten. Verlag Paul Parey, Berlin und Hamburg. P.: 72-75.
- StrliÄ, M., Mulej, I. (2000). OpraÅ¡evanje malo drugaÄe. Proteus, 63 (3): 104-108.